|
| ||||||
|
| |||||
|
Тема 2. ОТДЕЛ МОХООБРАЗНЫЕ (BRYOPHYTA)1. Общая характеристика Мохообразных. 2. Цикл развития Мохообразных. 3. Классификация Мохообразных. Класс печеночники, или печеночные мхи (Marchantiopsida, или Hepaticopsida). 4. Класс антоцеротовые (Anthocerotopsida). 5. Класс листостебельные мхи (Musci, или Bryopsida)
Общая характеристика Мохообразных
Мохообразные - это уникальная группа высших растений, которые характеризуются преобладанием в цикле воспроизведения гаплоидного гаметофита над диплоидным спорофитом. Мохообразные имеют длительную историю своего существования. Ранние палеонтологические находки датируются концом девона - началом карбона. Однако в отличие от других высших растений мохообразные ни по числу видов, ни по участию в сложении растительного покрова никогда не занимали на Земле господствующего положения. Это объясняется прежде всего тем, что гаметофит мохообразных составляет основное тело растения. Для осуществления полового процесса, который происходит у них с помощью сперматозоидов, необходима влажная среда. Следовательно, мохообразные вынуждены находиться в приземных, наиболее влажных слоях атмосферы, что и обусловливает их малые размеры. Кроме того, поскольку гаплоидный гаметофит обладает меньшим генетическим потенциалом по сравнению с диплоидным спорофитом, ему присущ менее активный процесс обмена веществ. Обладая небольшими размерами и будучи связанными с влажными местообитаниями, мохообразные с момента своего возникновения оказались под пологом более крупных растений, т.е. в условиях пониженного освещения. Поэтому смена эпох, изменения в растительном покрове земли не привнесли значительных изменений в условия жизни мохообразных. Это способствовало сохранению у них множества изначальных, т.е. примитивных, признаков. Вместе с тем за длительный период своего существования мохообразные по-разному адаптировались к условиям среды. Поэтому своеобразие мохообразных заключается прежде всего в том, что специализация их анатомо-морфологических структур и физиологических процессов осуществлялась на базе их низкого эволюционного уровня. К числу примитивных признаков мохообразных относится способность при неблагоприятных условиях переходить в состояние анабиоза, что отражает общую низкую активность обмена веществ. В состоянии анабиоза, при котором практически прекращаются все обменные процессы, растения могут длительное время (иногда десятилетиями) переносить экстремальные условия - дефицит влаги, высокие (до 70, 120 °С) или отрицательные температуры. При этом мохообразные не активно противостоят факторам среды, а пассивно «уходят» от неблагоприятных условий. Многие мохообразные относятся к числу олиготрофов, т.е. растений, способных произрастать на почвах, бедных минеральными веществами. Это свойство, по-видимому, также связано с их пониженным обменом веществ и незначительными размерами, но оно позволяет мохообразным осваивать наиболее бедные места обитания. Мохообразные насчитывают от 22 до 27 тыс. видов. Они широко распространены на всех материках земного шара, но имеют довольно ограниченную экологическую приуроченность. Большинство из них являются обитателями тенистых лесов, значительную роль они играют в формировании болот, встречаются на влажных лугах, по берегам водоемов.
Цикл развития Мохообразных
Индивидуальная жизнь мохообразных начинается с прорастания спор. Чаще всего при набухании споры экзина лопается, а интина вместе с содержимым споры вытягивается в виде сосочка, который, делясь, дает начало либо однорядной нити, либо однослойной (редко многослойной) пластинке, несущей ризоиды. Эта начальная стадия развития гаметофита называется стадией протонемы (от греч. protos - первичная и nema - нить). Важно подчеркнуть, что уже с самых первых этапов жизни молодой гаметофит целиком зависит от условий среды и при малейших неблагоприятных условиях обречен на гибель. Реже спора начинает делиться под оболочкой, и при разрыве экзины высвобождается уже многоклеточная протонема. Протонема часто подразделяется на хлоронему - зеленую ассимилирующую часть и ризонему - подземную бесцветную часть. В зависимости от принадлежности к той или иной систематической группе протонема либо постепенно превращается во взрослый талломный гаметофит (у печеночников), либо на протонеме формируются почки, дающие начало взрослому листостебельному гаметофиту (у листостебельных мхов). Общим для гаметофитов мохообразных является наличие лишь одной инициальной клетки, имеющей форму трехгранной пирамиды. Особенность анатомического строения талломных и листостебельных гаметофитов заключается в том, что эпидерма их лишена кутикулы и типичных устьиц, а проводящая система, даже самая совершенная, не имеет ни ситовидных клеток, ни трахеид. На гаметофите формируются одноклеточные или многоклеточные ризоиды. Несмотря на относительную простоту строения, мохообразные обладают самым сложноустроенным гаметофитом во всем растительном мире, что делает эту группу уникальной. На гаметофите формируются половые органы - архегонии и антеридии, как правило, располагающиеся на однорядных ножках. Архегонии возникают либо из точки роста, либо около нее и различными способами оказываются защищенными от высыхания. Оплодотворение возможно только при наличии воды. Из оплодотворенной зиготы развивается бесполое поколение - диплоидный спорофит, который всю жизнь проводит на гаметофите. Он либо частично, либо полностью питается за счет него. Сообразно функции и образу жизни спорофит всегда состоит из двух обязательных частей - гаустории и коробочки со спорами. После первого деления зиготы нижняя клетка формирует многоклеточную гаусторию, которая внедряется в ткань гаметофита. Она поглощает воду и питательные вещества, необходимые для развития спор. Из верхней клетки поделившейся зиготы формируется коробочка, которая либо непосредственно выполняет функцию спорангия (у печеночников), либо имеет сложное строение; в этом случае спорангий, как самостоятельный орган, развивается внутри коробочки (у мхов). У многих мохообразных в результате интеркалярного роста между коробочкой и гаусторией развивается ножка, выносящая коробочку наружу и способствующая лучшему рассеиванию спор. В силу своей специфичности спорофит мохообразных получил название спорогония.
Классификация Мохообразных
В основу классификации мохообразных бриологи кладут разные признаки, но во многом она определяется их взглядами на происхождение этой группы (проблема происхождения моховидных решается также неоднозначно). Согласно наиболее распространенной классификации, мохообразные подразделяются на три класса - печеночники (или маршанциевые), антоцеро-товые и листостебельные мхи (или просто мхи). В основу этой классификации положено морфологическое строение тела гаметофита, особенности строения ризоидов, строение и характер раскрывания коробочек, а также географическое распространение. Однако представители класса антоцеротовых характеризуются настолько отличающимся строением коробочки, что некоторые биологи придают им статус самостоятельного отдела.
Класс печеночники, или печеночные мхи (Marchantiopsida, или Hepaticopsida)
Гаметофиты печеночников, имеющие либо пластинчатые (слоевищные), либо листостебельные талломы, характеризуются дорзовентральным строением. У печеночников это определяется тем, что вентральная, или брюшная, сторона отличается от дорзальной, или спинной, стороны наличием ризоидов. У листостебельных печеночников дорзовентральность обусловлена тем, что листья располагаются в 2 ряда по бокам от оси, а на нижней - вентральной стороне формируются брюшные чешуйки - амфигастрии, которые отличаются от листьев и размерами, и формой, и характером развития. Листья печеночников в отличие от листьев мхов всегда однослойные и состоят из однородных клеток. Ризоиды их только одноклеточные, хотя могут иметь разное строение. Для спорогония печеночников наиболее характерно то, что в однослойной или многослойной коробочке споры развиваются непосредственно под стенкой, т.е. она функционально является спорангием. Стенка при созревании либо сгнивает, либо растрескивается, но она не имеет механизмов для активного выбрасывания спор. Функцию разрыхления спор выполняют особые вытянутые клетки - элатеры. Они имеют спиральные утолщения и, обладая гигроскопичностью, могут раскручиваться или закручиваться; поэтому иначе они называются пружинками. У некоторых видов вместо элатер образуются питательные клетки. Органы полового размножения размещаются на особых подставках и возвышаются над слоевищем. У мужских гаметофитов имеются подставки в виде восьми лопастных дисков, сидящих на ножке. На верхней стороне дисков находятся отверстия, ведущие в антеридии с двужгутиковыми сперматозоидами. На женских гаметофитах подставки имеют вид многолучевой звезды; между лучами подставки группами (шейкой вниз) расположены архегонии, прикрытые покрывальцем - перианцием. Имеется общее покрывальце перихеций. В дождливую погоду или с росой сперматозоиды попадают на женские подставки и проникают в архегонии. После оплодотворения из зиготы развивается спорангий в виде овальной коробочки, сидящей на очень короткой ножке. Внутри коробочки в результате мейоза образуются гаплоидные, но физиологически разные споры. К моменту созревания спор коробочки лопаются и споры высыпаются. Споры, выпадающие из спорангия, сначала дают пластинчатую слабо развитую протонему, из которой затем развивается новый (мужской или женский) гаметофит. У печеночников имеет место и вегетативное размножение, осуществляющееся выводковыми почками, образующимися в выводковых корзиночках на слоевищах (рис.2). Протонемы печеночников очень слабо развиты. У слоевищных видов они по форме мало отличаются от взрослого гаметофита. У всех исследованных видов печеночников обнаружена микориза, которая отсутствует у листостебельных мхов. Подавляющее число видов печеночников приурочено к влажным тропическим и субтропическим лесам, где они часто образуют сплошной напочвенный ковер, чехлом окутывают стволы деревьев; будучи эпифитами, оплетают листья мелких и крупных растений. Лишь относительно небольшое число видов печеночников распространено в умеренных областях Северного и Южного полушарий. Сочетание таких специфических признаков, как строение коробочки, дорзовентральность гаметофита, наличие микоризы, своеобразный ареал, говорит о том, что печеночники, возникнув одновременно со мхами, шли в своем развитии независимым, параллельным со мхами путем. Класс печеночники включает в себя два подкласса - маршанциевые и юнгерманиевые.
Рис. 2. Жизненный цикл печеночников (Маршанция)
Класс антоцеротовые (Anthocerotopsida)
Для представителей этого класса характерны пластинчатые талломы большей частью розетковидной формы. По краям талломов возникают меристематические клетки, которые формируют лопасти, налегающие друг на друга и придающие таллому курчавость. Талломы состоят из однородных тонкостенных клеток; верхние из них содержат хроматофоры с пиреноидами, которые резко отличаются от хлоропластов всех прочих высших растений. Наиболее широко распространен род антоцерос (Anthoceros), объединяющий около 200 видов, которые распространены в умеренных областях обоих полушарий и в тропиках. В России встречаются только 2 вида, которые произрастают на сырой почве. Их талломы размером 2-3 см, по краям однослойные, в центре многослойные; на их нижней стороне развиваются гладкие ризоиды и щелевидные отверстия, ведущие в полости. Эти полости сначала заполнены слизью, которая активно поглощает влагу; позже туда проникают нити ностока (Nostoc). На верхней поверхности таллома возникают пластинчатые или волосовидные придатки, создающие паутинистый налет, который также способен поглощать влагу из атмосферы и создавать вокруг слоевища влажную среду. В сухие годы у некоторых видов антоцероса на лопастях таллома могут формироваться клубневидные утолщения, покрытые сверху мертвыми опробковев-шими клетками; эти клубеньки способны переносить крайне неблагоприятные условия, а затем прорастать в новые растения. Антоцеросы однодомны, но антеридии созревают раньше архегониев, что предотвращает самооплодотворение. После оплодотворения развивается спорофит, который по своему внешнему и внутреннему строению значительно отличается от спорофитов прочих мохообразных. Спорогоний антоцероса состоит из гаустории и длинной узкой, слегка изогнутой коробочки роговидной формы. Между гаусторией и коробочкой находится меристематическая ткань, благодаря которой коробочка растет своим основанием до самой зимы. На верхушке коробочка вскрывается двумя длинными створками и высеивает зрелые споры, в то время как в основании ее близ меристемы находятся споры незрелые и только что возникшие. В центре коробочки проходит узкая колонка, которая также все время подрастает снизу. Стенка коробочки сверху покрыта эпидермой с типичными устьицами, а под эпидермой располагается фотосинтезирующая ткань. Особенность гаустории заключается в том, что иногда она прорастает сквозь пластинку гаметофита и внедряется в почву, развивая на своей поверхности ризоиды. Все это подтверждает взгляды о том, что когда то спорофит вел самостоятельный образ жизни и имел более сложное строение.
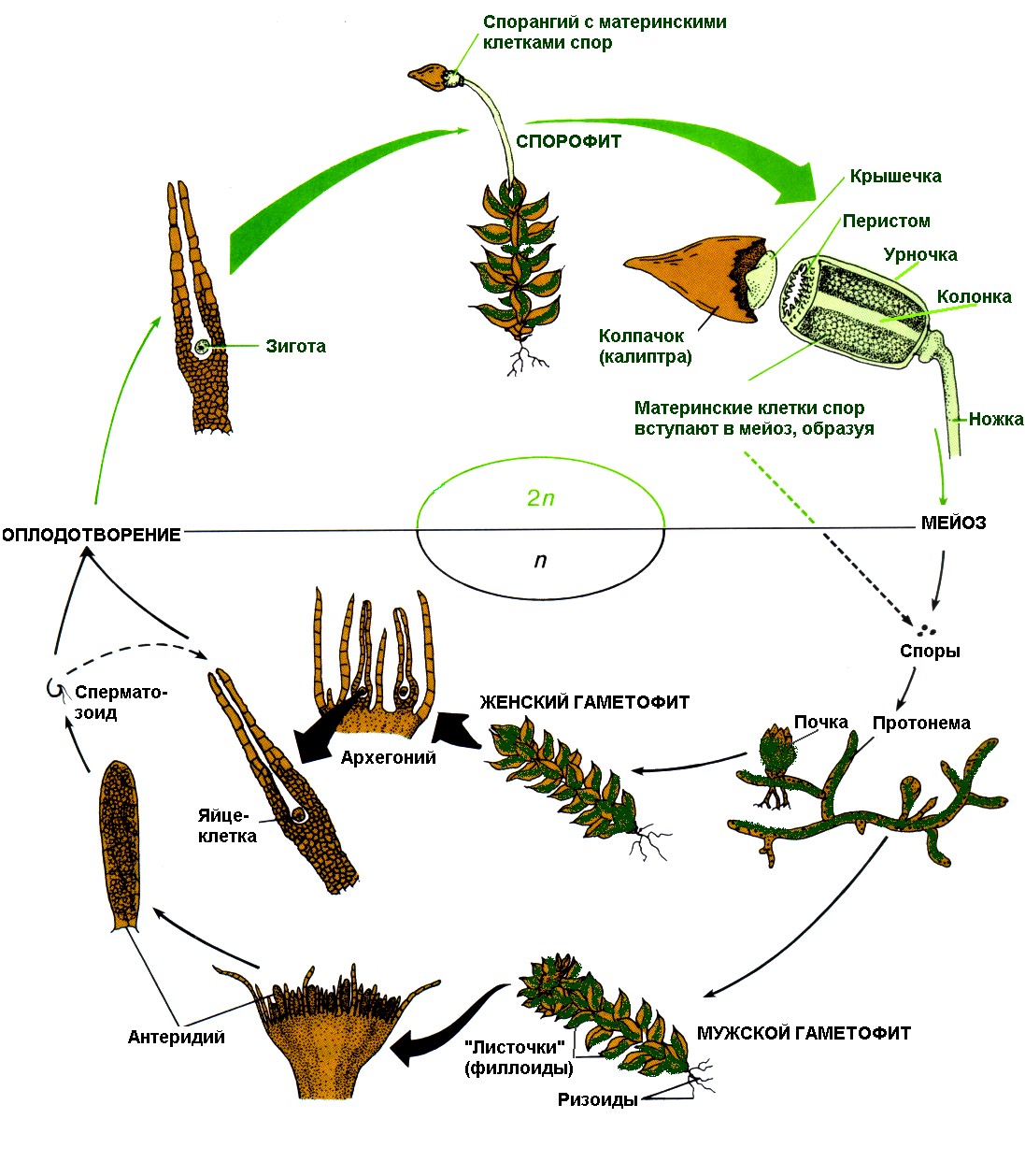
Класс листостебельные мхи (Musci, или Bryopsida)
Представители этого класса характеризуются наличием листостебельных побегов и, как правило, радиальной симметрии. У зеленых и андреевых мхов листья многослойные; у сфагнума они хотя и однослойные, но при этом дифференцированы на два типа высокоспециализированных клеток. Развивающие
Рис. 3. Сфагнум (Sphagnum): А — внешний вид растения (1— боковая веточка с листьями; 2 — стебель); Б — анатомическое строение листа (вид сверху): 1 — гиалиновая клетка; 2 — гиалиновое утолщение; 3 — пора; 4 — хлорофиллоносная клетка
ся на стеблях или листьях ризоиды всегда многоклеточные, часто обильно ветвящиеся. Они образуют войлок на поверхности стеблей, либо, переплетаясь, формируют шнуровидные тяжи. Коробочка спорогония в отличие от печеночников имеет сложное строение; она состоит из урночки и крышечки. Внутри урночки разными путями формируется спорангий со спорами. При созревании крышечка отскакивает, способствуя активному выбрасыванию спор, поэтому элатеры у мхов отсутствуют. Развивающаяся из споры протонема резко отличается от взрослого гаметофита; она имеет форму ветвящейся нити либо пластинки. Подавляющее число видов мхов в отличие от печеночников приурочено к сырым местообитаниям умеренных и холодных областей, где они играют большую роль в формировании ландшафтов северной тайги и тундры. В тропиках они распространены главным образом в горах; в степях и пустынях встречаются лишь одиночные виды мхов. Незначительное число видов перешло ко вторично водному образу жизни. Несмотря на широкое распространение мхов по всему земному шару, каждый род, а иногда вид, проявляет избирательность к механическому и химическому составу почв; одни мхи, предпочитающие богатые почвы, селятся на гниющих стволах, на навозе, на трупах. Другие, а их большинство, являются олиготрофами - они осваивают бедные песчаные или каменистые почвы. Класс мхи включает более 14,5 тыс. видов и объединяет 3 подкласса - сфагновые, зеленые и андреевы мхи. Подкласс сфагновые, белые, или торфяные мхи (Sphagnidae). Этот подкласс представлен одним порядком Sphagnales, одним семейством Sphagnaceae с одним родом сфагнум (Sphagnum), объединяющим свыше 300 видов, которые морфологически довольно трудно различаются. В России произрастает 42 вида. Все представители беловато-зеленого цвета, не имеют ризоидов. От стебля растения отходят боковые веточки, на верхушке собранные в головку (рис.3, А). Побеги сфагнума усажены мелкими листьями. Листья однослойные, состоят из двух типов клеток - хлорофиллоносных и мертвых водоносных (гиалиновых). За счет наличия в листе мертвых гиалиновых клеток, способных удерживать воду, они обладают гигроскопическим свойством. Фотосинтез протекает в живых ассимиляционных клетках, содержащих хлоропласты (рис.3, Б). При высыхании мертвые клетки заполняются воздухом, и мох становится беловатым, за счет чего и получил название белый мох. Сфагновые мхи наиболее широко распространены в умеренной зоне Северного полушария, где на обширных пространствах они формируют верховые сфагновые болота. Поселяясь на влажных местах, многие виды сфагнума способствуют быстрому заболачиванию этих территорий благодаря способности активно и быстро поглощать влагу и прочно ее удерживать. Это объясняется высокой специализацией морфологического и анатомического строения гаметофита. Развитие гаметофита начинается с прорастания споры, которая вначале формирует однорядную нить. Если прорастание споры происходит в воде, то сохраняется нитчатая структура. При прорастании ее на поверхности влажной почвы сначала из нити образуется округлая, а затем лентовидно-лопастная пластинка размером до 1 см с многоклеточными ветвящимися ризоидами. По краям пластинки формируются почки, из которых развиваются олиственные побеги гаметофита. Молодые побеги, возникающие на протонеме, густо покрыты мелкими листьями, а в основании развиваются многоклеточные ризоиды. С началом ветвления побегов образование ризоидов прекращается, и взрослые растения сфагнума полностью лишены их. Половые органы развиваются на специализированных боковых генеративных побегах, расположенных у вершины главного побега в пучках вместе с вегетативными побегами. Созревание половых органов происходит осенью, а половой процесс может происходить либо осенью, либо весной следующего года. Одни виды сфагнума однодомные, другие - двудомные. Сфагновые мхи демонстрируют высочайшую степень специализации гаметофита, обеспечивающую поглощение воды как из почвы, так и из атмосферы, быстрое ее передвижение по всему телу гаметофита и прочное удержание воды растением. Однако принцип поглощения воды растением основан исключительно на физических законах - капиллярности, гигроскопичности, набухании, т.е. так же, как у юнгерманниевых, имеет место тонкая специализация на базе низкого эволюционного уровня. Подкласс андреевые мхи (andreaeidae). Андреевые мхи - это небольшая группа мхов, представленная одним семейством Andreaeaceae с единственным родом андреа (Andreaea), или андреев мох, объединяющим 120 видов. Андреевые мхи распространены в умеренных и холодных областях Северного и Южного полушарий, довольно высоко поднимаются в горы и приурочены в основном к скалам и камням. По анатомическому и морфологическому строению они четко отличаются от сфагновых и зеленых мхов. 1) Развитие гаметофита начинается с прорастания бесцветных спор, которые делятся под оболочкой, и уже многоклеточные клубневидные тельца разрывают экзину и прорастают в зеленую лентовидную протонему. 2) Однослойные листья с однородными клетками способны к длительному нарастанию своей вершиной, в результате чего формируются бесцветные гигроскопические волоски, способные поглощать влагу из атмосферы. 3) Стебли лишены проводящих пучков. 4) Спорогоний состоит из коробочки и гаустории. Перед созреванием коробочки развивается ложная ножка, выносящая коробочку вверх. 5) Коробочка без крышечки растрескивается четырьмя продольными щелями, образующимися в средней части коробочки. При подсыхании стенки коробочки дугообразно изгибаются, что приводит к увеличению просвета щелей, через которые высыпаются споры. Подкласс зеленые мхи (Bryidae). К числу особенностей этого подкласса относятся: 1) наличие хорошо развитой нитчатой протонемы, на которой формируются побеги; 2) наличие в листе средней жилки, даже если листовая пластинка однослойная; 3) сложное строение спорогония, состоящего из коробочки, ножки и гаустории; 4) наличие в коробочке перистома и колонки, доходящей до вершины. Характерным представителем подкласса является мох кукушкин лен обыкновенный (Polytrchum commune). Кукушкин лен - один из наиболее высокорослых мхов, его стебель достигает 50 см в высоту. Произрастает на сырой почве в лесах и болотах, образуя крупные подушкообразные дернины. Стебель мха - прямостоячий, неветвящийся, густо покрыт жесткими линейно-шиловидными листьями. Листья многослойные, в отличие от других мхов состоящие из однородных рядов хлорофиллоносной ткани. Подземная часть представлена многолетними ветвящимися ризоидами. Кукушкин лен относится к двудомным растениям. На женском растении (гаметофите) между верхними салатового цвета листьями образуются архегонии - женские половые органы. Архегоний - многоклеточное образование колбовидной формы. Суженная часть - шейка, а расширенная часть - брюшко, в котором формируется крупная яйцеклетка. На мужском растении (гаметофите) среди верхних красных листьев развиваются антеридии - мужские половые органы, в которых образуются двужгутиковые сперматозоиды. Антеридии имеют вид продолговатых или округлых мешочков на ножке. При созревании архегония шейковые, или канальцевые, клетки ослизняются, и на их месте формируется узкий канал, по которому сперматозоид может проникнуть в яйцеклетку. В период обильных дождей или снеготаяния сперматозоиды подплывают к архегониям. Как предполагают, они обладают хемотаксисом к содержимому слизи архегония. Один из сперматозоидов проникает в архегоний и продолжает движение к яйцеклетке. Слияние гамет и дальнейшее развитие зиготы происходит внутри архегония. Через несколько месяцев из зиготы вырастает спорофит (спорогон) в виде коробочки на длинной ножке.
Рис.4. Жизненный цикл листостебельных мхов (Кукушкин лен)
Нижняя часть ножки превращена в гаусторий (присоску), внедряющийся в тело женского гаметофита. Спорофит, таким образом, лишен самостоятельности и полностью зависит от гаметофита. Сверху коробочка прикрыта легко опадающим колпачком (остаток архегония) с тонкими, направленными вниз волосками, напоминающими льняную пряжу (отсюда и название растения). Внутри коробочки - в спорангии - происходит образование спор мейозом. При созревании спор колпачок, а затем и крышечка отделяются, и споры высыпаются из отверстий наверху коробочки (урны). Коробочка имеет ряд зубцов - перистом, прикрывающих отверстия во влажную погоду. Одинаковые по форме споры (изоспоры) разносятся ветром, а затем попадают в почву и прорастают в зеленую нить - протонему (типа нитчатой водоросли), на которой из почек формируются листостебельные побеги. Таким образом, эти побеги с протонемой представляют гаметофит, имеющий гаплоидный набор хромосом. На это цикл замыкается, и все повторяется снова (рис.4). Как показывает обзор отдела мохообразных, эту группу объединяет много общих черт, прежде всего преобладание в жизненном цикле гаметофитного поколения. На происхождение этого своеобразного жизненного цикла существует две группы воззрений. Согласно широко распространенному мнению, предковые формы всех высших растений обладали изоморфной сменой поколений, каждое из которых вело самостоятельный образ жизни. В дальнейшем мохообразные пошли по пути сокращения спорофита, что привело к потере его самостоятельности. На это указывает наличие в спорогонии некоторых мохообразных редуцированных устьиц, проводящих тканей в колонке и ножке мхов, случаи прорастания гаустории сквозь ткань гаметофита в почву и развитие на ее поверхности ризоидов. В результате значительной редукции целое поколение свелось до органа, производящего споры. Напротив, половое поколение оказалось в жизненном цикле ведущим; хотя оно и дало большое многообразие жизненных форм, но никогда не смогло занять господствующего положения на Земле. Согласно другим представлениям, начало мохообразным, как и всем прочим архегониальным растениям, дали зеленые водоросли порядка хетофоровых. Они обладали гетеротри-хиальными талломами и самыми разнообразными типами циклов развития. Начало одним мохообразным дали водоросли, обладавшие изоморфной сменой поколений с последующей редукцией спорофита. Другие же мохообразные с самого начала унаследовали цикл развития с преобладанием гаметофита. | ||||||
|
| ||||||
| Сайт создан по технологии «Конструктор сайтов e-Publish» | ||||||